A halofita növények sókiválasztása
A sótűrő – szaknyelven halofita – növények a szárazföldi virágos növények egy ökológiai alapon jól elkülönült csoportját képviselik. Tagjaik hatékonyan alkalmazkodtak a sóban gazdag élőhelyekhez: a fél-sivatagokhoz, a sós mocsarakhoz és a tengerpartokhoz. Ezeken az élőhelyeken a néha extrém magas sótartalom feltételei mellett is képesek a fajra jellemző módon növekedni , fejlődni, szaporodni, fennmaradni.
Sós élőhelyről általában akkor beszélünk, ha a talajban a NaCl-koncentráció 0,5% vagy ezt meghaladó érték (a tengervíz NaCl-tartalma 3-5%, kb. 0,5 mol/l). A magas sótartalom egyrészt a növények számára stresszt okozó toxikus ionhatás, másrészt a közvetlen környezet ozmotikus potenciáljának növelésével csökkenti a növények számára hozzáférhető víz mennyiségét Emellett a talaj magas nátriumion-koncentrációja rontja a talaj fizikai tulajdonságait pl tömöríti a talajszerkezetet, csökkenti a kicserélhető tápelemek mennyiségét, a felvehető víz tartalmat stb.
Számos halofita növény a levélfelületén lévő speciális sókiválasztó rendszer segítségével sajátos sókiválasztással szabadul meg a felesleges és toxikus hatású konyhasó/ nátrium és klorid/ ionoktól. A klasszikus mirigyekkel történő un exogén kiválasztással szemben itt egy másféle elválasztásról beszélünk, melyet a szekréciótól eltérően újabban rekréciónak neveznek; ekkor a növény úgy választja ki az anyagokat, ahogyan felvette azokat.
Az egyes halofiták sókiválasztó képződményei a faj rendszertani helyzetétől függően eltérő felépítésűek, működésük lényege azonban hasonló: a folyamat kezdetén a kiválasztó sejtbe irányuló, jelentős mennyiségű anyagcsere energiát/ATP/ igénylő, aktív nátrium iontranszport megnöveli a sejt ozmotikus értékét és csökkenti a vízpotenciálját. Ennek következtében egy sejtbe történő passzív vízmozgás jön létre. A sejtmembrán vízvezető csatornáin/akvaporin/ megvalósuló gyors vízbeáramlás miatt a kiválasztó sejtekben megnő a belső hidrosztatikus nyomás/turgor/, amely hajtóerőként működik és kifelé nyomja a vizet és a benne oldott anyagokat( jelen esetben elsősorban a konyhasó ionokat). A kiválasztó képződmény oldalában található sajátos sejtfalvastagodások és az ide berakódott paraszövet megakadályozzák az oldalirányú, vagy visszafelé történő vízkiáramlást. A sóoldat így a legkisebb ellenállást mutató felszín irányába mozog és megtörténik a levél felszínén a kiválasztás.
A felületi sókiválasztás legegyszerűbb képviselői a mindössze két-három sejtből álló a levelek bőrszövetében fejlődő, általában szabad szemmel is megfigyelhető különböző nagyságú világos gömböcskék az un hólyagszőrök. Az epidermiszből kiemelkedő hólyagok többnyire szürkés színű enyhén érdes, lisztes tapintású bevonatot képeznek a levélfelszínen. Ezek a speciális, néhány sejtből álló módosult epidermális szőrök/trichomák/ sókiválasztó/sóleadó szerepet látnak el és a hazai flórában is találkozhatunk velük a libatopfélék/Chenopodiaceae/ családban.
Egyszerű esetben/ pl kristályvirág féléknél/ az epidermisz sejtrétegben elhelyezkedő alapi/bazális/ sejtből és egy hozzá kapcsolt a felszínből kiemelkedő gömbszerű feji sejtből állnak. A laboda/Atriplex/ fajoknál gyakran a két sejt között egy közvetítő harmadik sejt/un nyaki sejt/ is található/1.ábra/ 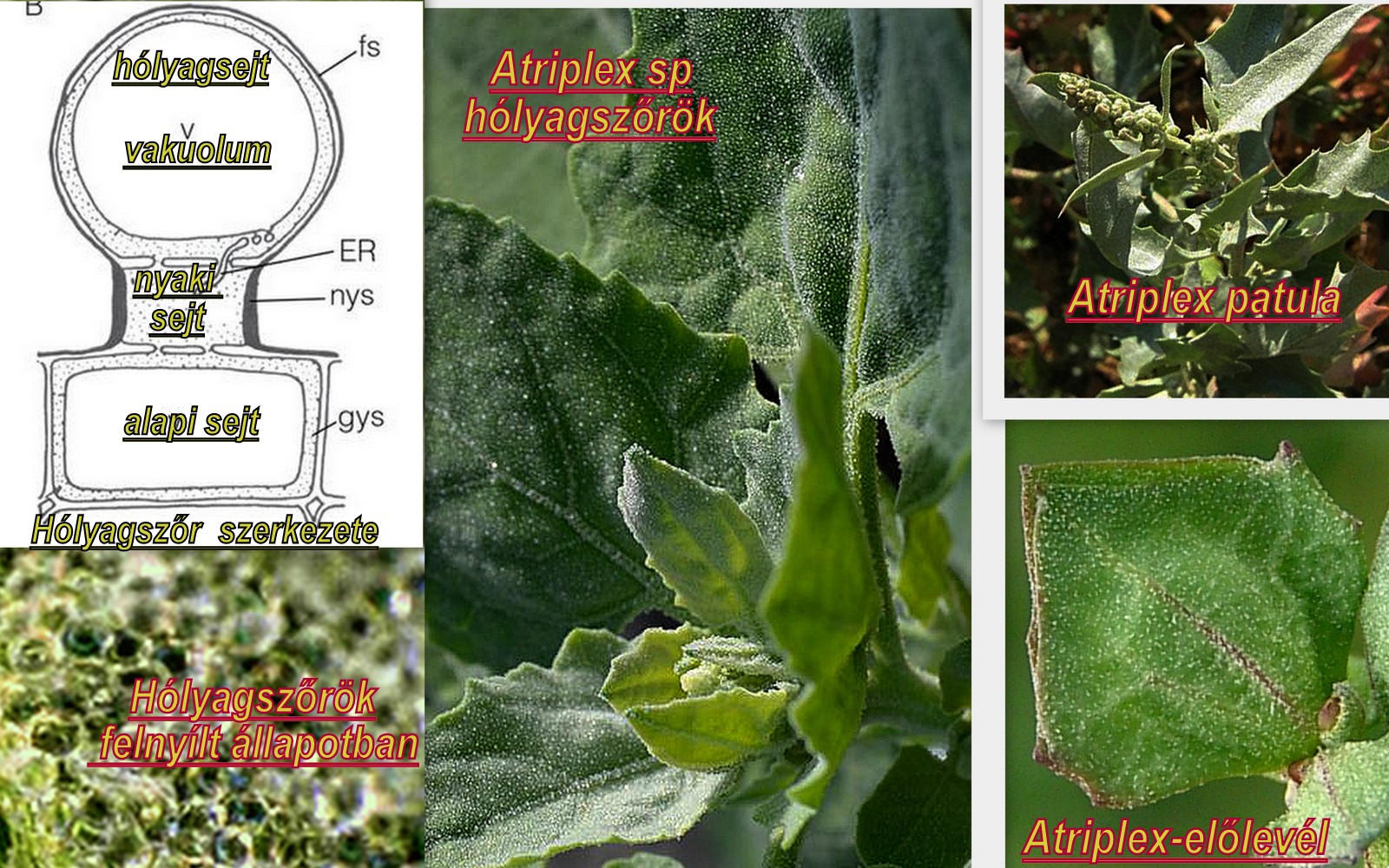
A felső hólyagsejt térfogatának nagy részét hatalmas központi helyzetű vakuolum foglalja el, amelyben a nátrium klorid kiválasztás és raktározás történik. Az alapi és a nyélsejtek élénk, intenzív anyagcseréjűek, belső membránokban és mitochondriumokban gazdagok, sejtfalaikban számos gödörkés vastagodást találunk, melyek a sejtek közötti transzportban fontos szerepet játszanak.
A nátrium és klorid ionok kivonása a hólyagsejt nagy centrális vakuolumát határoló membránban/tonoplaszt/ történik. Itt találjuk azokat a speciális, szelektív nátrium ion-szállító csatornákat, amelyek jelentős mennyiségű ATP felhasználásával gyűjtik be a nátrium ionokat a vakuolumba. A nátrium felvétel tehát membrán ATP-ázok által közvetített aktív transzport folyamat, lényegében egy nátrium ion- proton ioncsere. A klorid anionok felvétele külön úton más mechanizmussal történik és nem igényel közvetlen ATP felhasználást. A töltésegyensúlyt ugyanakkor ez utóbbi folyamat biztosítja. Az ionakkumuláció természetesen növeli a hólyagsejtek ozmotikus értékét, amely egy gyors passzív vízfelvételt eredményez. A belső hidrosztatikus nyomás növekedése pedig gyakran vezet a hólyagsejt szétszakadásához. A felszínre jutott sóoldat beszáradva kristályokat képez, esetleg a csapadék mossa le a növényről. Gyakran előfordul, hogy a sóval telt hólyagszőr felrepedése előtt a nyaki részen lehajlik, később letörik és lehull a talajra.
A bonyolultabb felépítésű sómirigyek rendszerint a bőrszövetbe mélyednek, egy epidermisz gödör mélyén ülnek. A kiválasztott sócsepp nem terül szét a felszínen, az oldat vize fokozatosan elpárolog, a sókristályok kiválnak és a folyamatos kiválasztás/rekréció/ következtében fokozatosan gyarapodnak. Egy kiadós eső, vagy a dagály hullámai mossák le a sót időnként a levélfelszínről.
A sómirigyek legegyszerűbb típusát az atlanti partok sós mocsaraiban tömeges, nálunk kedvelt díszfűként ültetett Spartina/Cordgrass-Zsinegfű/ fajok levelein figyelhetjük meg. A sókiválasztó rendszer felépítése ezekben a fajokban a más növényekben vizet kiválasztó hidatodákhoz hasonlít. A sóvirág/Limonium/ és a mangrove fajok/pl Avicennia/ összetett felépítésű sómirigyei adják általában az iskolapéldát a növényi sókiválasztás bemutatásához.
A több sejtből álló kiválasztó rendszer alsó-oldalsó/bazális-laterális/ részén találjuk az un gyűjtősejteket. Ezekből a sejtfal plazmodezmákon át citoplazma hidakon un szinplaszt transzporttal jut az oldat a központi helyzetben álló hengeres, sugarasan elrendezett tényleges kiválasztó sejtekbe/un szekréciós sejtek-sugársejtek/. Az egyirányú oldat kiáramlást, az oldalirányú, esetleg visszafelé történő vízmozgást a rendszert oldalról és alulról övező vastagodott, parásodott sejtfalak vízgátként biztosítják. A sómirigyeket gyakran felül egy pórusokkal átlyuggatott, fordított tányérhoz hasonlítható kuti- kula sapka is védi. A tényleges kiválasztás a pórusokon keresztül történik. Több fajnál nem sikerült kimutatni ezt a perforációt, ilyenkor a kuti-kula lemez és a szekréciós sejtek falai között halmozódik a kiválasztott anyag, és a fedőréteg felrepedésével kerül a szabadba/2.ábra/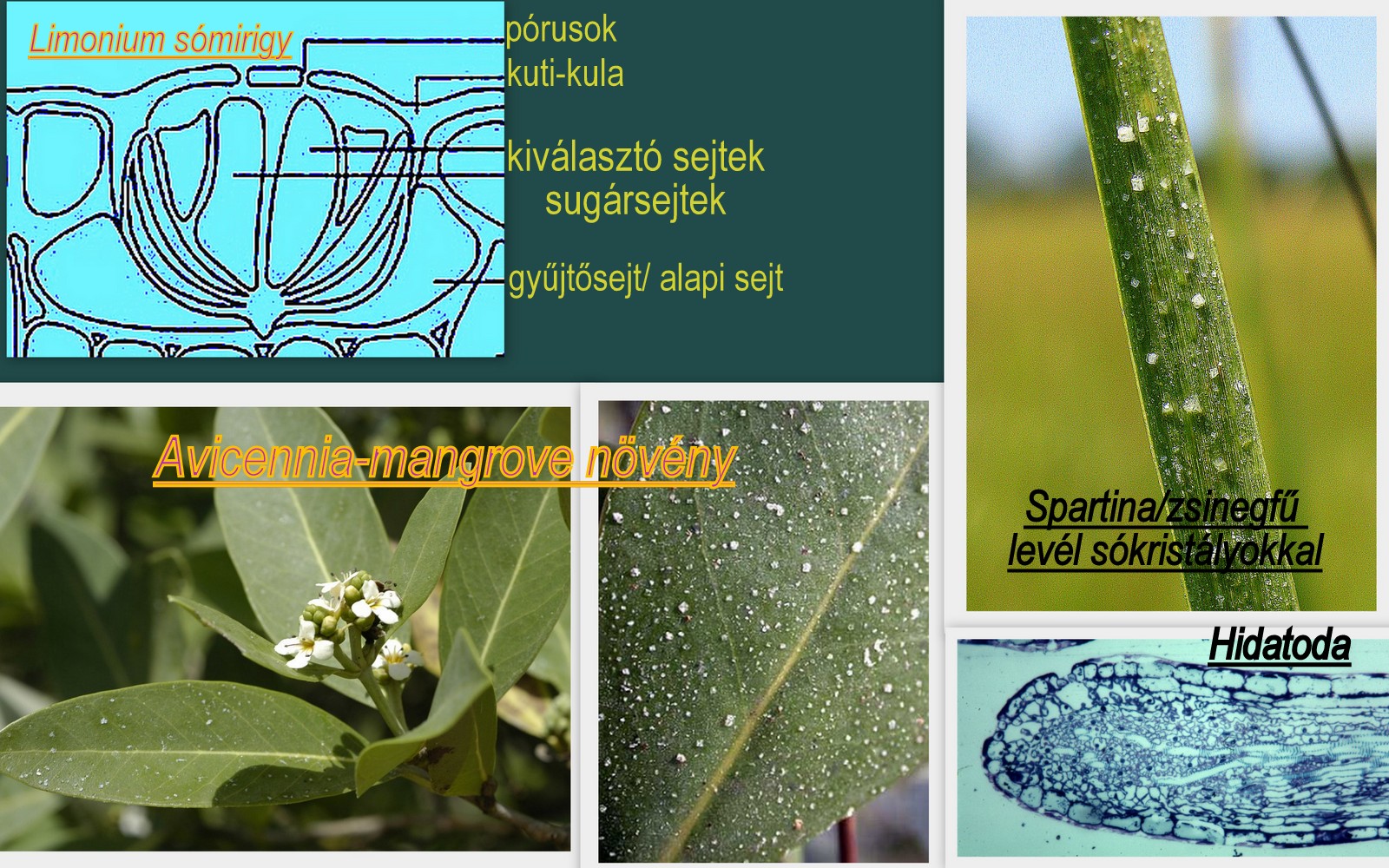
A sókiválasztás – elsősorban a jelentős nátrium iontranszport miatt – rendkívül energia igényes ugyanakkor azonban hatékony folyamat. A Tamarix fajoknál például a számítások szerint egy mol konyhasó kiválasztásához mintegy 20-25 mol ATP szükséges. Az Atriplex fajok a felvett sóionok több mint 80%-át ki tudják választani a hólyagszőrökön keresztül.
A sómirigyek száma is nagyon eltérő lehet a levelekben. Míg a sóvirágnak/Limonium/ 3000 sómirigye lehet cm²-enként, addig a pázsitszegfűnél (Armeria maritima) csupán 590 sómirigyet, a bagolyfűnél (Glaux maritima) 800 sómirigyet számoltak össze egy cm² felületen.
A sómirigyek lehetővé teszik számos halofiton, különösen a sós élőhelyeken előforduló nem szukkulens halofitonok – pl. Plumbaginaceae- (Limonium spp., Statice spp.) és Tamarix-fajok vagy mangrove növények(pl Avicennia)– számára, hogy aktívan megszabaduljanak a felesleges sótól, és ezáltal megakadályozzák a nagy mennyiségű és toxikus sókoncentrációk kialakulását a szövetekben.
